
Review
Colonization and diversification of Gala
´
pagos
terrestrial fauna: a phylogenetic and
biogeographical synthesis
Christine E. Parent
1,
*
, Adalgisa Caccone
2,3
and Kenneth Petren
4
1
Department of Biological Sciences, Simon Fraser University, 8888 University Drive,
Burnaby, Canada BC V5A 1S6
2
Department of Ecology and Evolutionary Biology, and
3
Yale Institute of Biospheric Studies, Yale University,
New Haven, CT 06520-8106, USA
4
Department of Biological Sciences, University of Cincinnati, Cincinnati, OH 45221-0006, USA
Remote oceanic islands have long been recognized as natural models for the study of evolutionary
processes involved in diversification. Their remoteness provides opportunities for isolation and
divergence of populations, which make islands remarkable settings for the study of diversification.
Groups of islands may share a relatively similar geological history and comparable climate, but their
inhabitants experience subtly different environments and have distinct evolutionary histories, offering
the potential for comparative studies. A range of organisms have colonized the Gala
´
pagos Islands, and
various lineages have radiated throughout the archipelago to form unique assemblages. This review
pays particular attention to molecular phylogenetic studies of Gala
´
pagos terrestrial fauna. We find
that most of the Gala
´
pagos terrestrial fauna have diversified in parallel to the geological formation of
the islands. Lineages have occasionally diversified within islands, and the clearest cases occur in taxa
with very low vagility and on large islands with diverse habitats. Ecology and habitat specialization
appear to be critical in speciation both within and between islands. Although the number of
phylogenetic studies is continuously increasing, studies of natural history, ecology, evolution and
behaviour are essential to completely reveal how diversification proceeded on these islands.
Keywords: adaptive radiation; diversification; evolution; phylogeny; phylogeography; speciation
The distribution of the tenants of this archipelago
would not be nearly so wonderful, if, for instance, one
island had a mocking-thrush, and a second island some
other quite distinct genus [.]. But it is the circum-
stance, that several of the islands possess their own
species of the tortoise, mocking-thrush, finches, and
numerous plants, these species having the same general
habits, occupying analogous situations, and obviously
filling the same place in the natural economy of this
archipelago, that strikes me with wonder.
(Darwin 1845)
1. INTRODUCTION
Islands have always attracted professional and amateur
students of biology alike. By their very nature, islands
are biologically simpler than continental regions and
therefore provide ideal geographical and historical
settings for the study of colonization and diversification
of species. If the timing of their geological formation is
known, such study can be conducted within a defined
time frame. Moreover, if an island is part of an
archipelago, then each island represents a replicate
natural experiment and therefore provides increased
statistical power to test ideas related to patterns
observed and processes inferred about diversification.
The Gala
´
pagos archipelago occupies a unique
position in the history of evolutionary studies, mainly
owing to its importance as a conceptual landmark
(Darwin 1859) and because it continues to shape our
understanding of evolutionary biology (Grant & Grant
2008). Notwithstanding its tropical climate, the Gala
´
-
pagos archipelago has been the stage of surprisingly few
animal diversifications compared with other Pacific
tropical island groups. With only one known exception,
evolutionary radiations on Gala
´
pagos comprise fewer
than 20 species, and include mainly lineages of land
birds, reptiles and terrestrial invertebrates. Among
vertebrates, the absence of amphibians and the virtual
absence of mammals are particularly striking and nearly
unique among terrestrial island ecosystems.
The goal of this review is threefold. It is (i) to review
literature on Gala
´
pagos animal groups that have
speciated on the island, (ii) to synthesize the available
evidence in light of classic hypotheses related to
colonization and diversification on islands, and (iii) to
suggest future avenues in the study of Gala
´
pagos faunal
diversification and how they can be used to aid in
conservation efforts.
Phil. Trans. R. Soc. B (2008) 363, 3347–3361
doi:10.1098/rstb.2008.0118
Published online 4 September 2008
One contribution of 15 to a Theme Issue ‘Evolution on Pacific
islands: Darwin’s legacy’.
* Author for correspondence ([email protected]).
3347 This journal is q 2008 The Royal Society

2. THE GALA
´
PAGOS ARCHIPELAGO
(a) Geographical setting
The Gala
´
pagos Islands are located in the Pacific Ocean,
approximately 960 km west of the South American
coast, straddling the equator at the 90th meridian west
(figure 1a). The Gala
´
pagos archipelago is composed of
13 major islands larger than 10 km
2
, 6 smaller islands,
over 40 islets with official names and many smaller
unnamed islets and rocks, for a total of approximately
8000 km
2
of land spread over 45 000 km
2
of water
(Snell et al. 1996). Of these islands, Isabela is the
largest (more than the total area of all the other islands
combined) with an area of 4588 km
2
and the highest,
with a maximum elevation of just over 1700 m. Isabela
is formed by six volcanoes that are interconnected
mostly by barren lava flows, which may represent a
geographical barrier to dispersal analogous to open
water for terrestrial animal species with low dispersal
capacity such as flightless terrestrial invertebrates.
(b) Geological history
The Gala
´
pagos Islands are a young, oceanic and
volcanic archipelago. Their geological history is rela-
tively well understood ( Nordlie 1973; Swanson et al.
1974; Bailey 1976; Cox 1983; Hall 1983; White et al.
1993; Geist 1996), with individual islands being
formed as the Nazca plate moved over a hot spot.
However, in contrast to the roughly linear arrangement
by age of the Hawaiian Islands (Price & Clague 2002),
the Gala
´
pagos Islands are clustered into groups of
similar age ( White et al. 1993). The age of the islands
increases moving eastward along the plate, with the
oldest islands located towards the southeast of the
archipelago. K–Ar age determinations and marine
fossils indicate a maximum age of the oldest land of
the order of 3 Ma, whereas geological plate motion
models set a maximum age of emergence around 4 Ma,
depending on the velocity of the Nazca plate
(Hickman & Lipps 1985; White et al. 1993; Geist
1996; D. Geist 2005–2008, unpublished data; figure 1a).
The discovery of drowned seamounts east of San
Cristobal extends the time the Gala
´
pagos have been
available for colonization to at least 14 Ma (Werner et al.
1999). However, the history of island production over
this hot spot could extend back 80–90 Ma, which is the
estimated age of the birth of the Gala
´
pagos hot spot
(Christie et al. 1992; Hoernle et al. 2002).
(c) Biogeography and ecology
The climate of the Gala
´
pagos Islands is unusually dry
for the tropics and has marked seasonality. Since the
archipelago is so isolated, ocean and wind currents
mostly influence its climate. There are two main
seasons: the warm season, typically January through
May, and the cool season from June to December. The
warm season is caused by warm ocean currents
sweeping southward from the direction of Panama,
which cause both sea and air temperatures around the
islands to rise. During this time the skies are normally
clear, with occasional heavy showers. During the cool
season, the weather is influenced by the Humboldt
Current, which brings cold water north from the
Antarctic along the west coast of South America and
then westward through the archipelago, which results
in cooler air temperatures. During the cool season, the
skies are usually overcast, but with little precipitation in
the lowlands. However, many parts of the highlands are
constantly wet at this time owing to a mixture of light
rain and mist. The higher islands have increasingly more
rain at progressively higher elevations and a humid
forest zone exists at 300–600 m on their windward
(eastern) sides. Precipitation is variable in occurrence
and quantity, even in the wet season. The direction of
the wind and oceanic currents changes in response to a
seasonal north–south shift in the Intertropical Conver-
gence Zone (ICZ). On an irregular basis, but typically
once every 3–6 years (Allan et al. 1996), this change in
direction becomes more dramatic and warmer and
wetter seasons occur with potentially 10 times more
than the normal annual rainfall (referred to as El Nin
˜
o
events). In the cool season during these years, the ICZ
moves greatly to the north of the islands, and the
archipelago falls under the influence of the cool, dry
wind and cool ocean currents (termed La Nin
˜
a events).
These cyclic changes in climate can have a pronoun-
ced effect on Gala
´
pagos vegetation (McMullen 1999)
and fauna (Grant & Grant 1993; Grant et al. 2004;
Steinfartz et al. 2007).
Vegetation on Gala
´
pagos can be separated into six
(or seven) altitudinal zones (Wiggins & Porter 1971;
van der Werff 1979) and the plant species composition
of each zone is a reflection of the humidity level of the
zone, with moisture level increasing with elevation
(McMullen 1999). The Gala
´
pagos flora comprises 749
angiosperm species, of which 216 are endemic to the
archipelago (just under 30%), and 7 of the 192 genera
of flowering plants listed by McMullen (1999) are
endemic to the islands.
(d) E ndemism and diversification
Like that of many other isolated oceanic islands, the
Gala
´
pagos fauna is impoverished and taxonomically
unbalanced when compared with Neotropical source
areas. Owing to their isolation and arid climate, the
Gala
´
pagos Islands have been successfully colonized by
only a subset of the diverse flora and fauna of the closest
continent. Some lineages that have colonized the
Gala
´
pagos archipelago have not differentiated from
their continental ancestor (and thus are considered
native) or have differentiated without radiating into
multiple lineages. Both of these scenarios apply to all
sea birds, more than one-third of land bird diversity,
and the great majority of terrestrial invertebrates
(Peck 2001, 2006).
Although relatively few lineages of Gala
´
pagos
terrestrial fauna have diversified within the archipelago
(associated with adaptation or not), the resulting
species constitute a large proportion of the terrestrial
vertebrate fauna of the islands (table 1). Bats (two
species) and rats are the only terrestrial mammals that
have naturally colonized the islands, and among
them only the rice rats have diversified within the
archipelago with 10 recognized species (three extant)
representing three lineages (Clark 1984). Amphibians
have not naturally colonized the islands. Seven
reptile lineages have colonized the Gala
´
pagos and
at least four diversified after their arrival, such that
34 of the 37 recognized reptile species are part of
3348 C. E. Parent et al. Review. Diversification of Gala
´
pagos terrestrial fauna
Phil. Trans. R. Soc. B (2008)
multispecies lineages. Of the 30 land bird species,
25 are endemic and 60% of the endemic species belong
to two lineages that have speciated within the
archipelago: Darwin’s finches (14 species) and Gala
´
-
pagos mockingbirds (4 species). By contrast, the
Gala
´
pagos Islands have approximately 1500 species of
native insects of which only approximately 47% are
endemic. Very few terrestrial insect lineages have led to
multiple species: only approximately 5% of the
approximately 1000 insect genera that have success-
fully colonized the islands have diversified (Peck 2006)
and those that have diversified have given rise to
lineages of 20 species or fewer. With 71 described
species, the radiation of Gala
´
pagos bulimulid land
Darwin (0.4–?)
Wolf (1.0–?)
v. Ecuador
v. Wolf
v. Darwin
v. Alcedo
v. Sierra Negra
v. Cerro Azul
Seymour Norte
Baltra
Plaza Sur
Santa Fe
(2.9–2.9)
Santa Cruz
(1.1–2.3)
Isabela (0.5–0.8)
Floreana (1.5–2.3)
San Cristobal (2.4–4.0)
Española (3.0–3.5)
50 km
PinzÓn
(1.3–2.7)
Santiago (0.8–1.4)
Rabida
(1.3–1.6)
Fernandina
(0.035– 0.07)
Pinta (0.7–?)
Marchena
(0.6–?)
Genovesa
(0.3–?)
Cocos
Galapagos Islands
Ecuador
1°
1°
91° 90° 89°
0°
(a)
98
99
96
95
99
100
100
95
100
100
100
83
83
92
100
82
93
92
100
100
100
96
77
abingdoni
Pinta
Española
San Cristóbal
Santa Cruz (Cerro Fatal)
Santa Cruz (Cerro Monturra)
Isabela
Isabela
Santiago
Pinzón
hoodensis
chatamensis
porteri
becki1
becki2
darwini
ephippium
guntheri1
guntheri2
vicina1
vicina2
vicina3
vicina4
vicina5
vandenburghi
microphyes
porteri
Santa Cruz (La Caseta)
0.005 substitutions/site
undescribed*
Floreana
guntheri3
guntheri4
chatamensis*
chilensis
carbonania
denticulata
(b)
B. quitensis
B. sp. 2 (Alcedo, Isabela)
B. sp. 3 (Alcedo, Isabela)
B. tanneri (Wolf, Isabela)
B. darwini (Santiago)
B. blombergi (Santa Cruz)
B. olla (santiago)
B. Perrus 1 (Fernandina)
B. sculpturatus (Santiago)
B. jacobi (Santiago)
B.sp. 9 (Santa Cruz)
B.sp. 6 (Sierra Negra,Isabela)
B.sp.7 (Sierra Negra,Isabela)
B.reibischi (santa Cruz)
B.sp. 8 (Alcedo, Isabela)
B.perrus 2 (Fernandina)
B. tortuganus (Darwin, Isabela)
B. ochsneri (Santa Cruz)
B. wolfi (Santa Cruz)
B. pinzonopsis (Pinzon)
B. indefatigabilis (Santiago)
B. nesioticus (Santiago)
B. sculpturatus (Santiago)
B. akamatus (Santa Cruz)
B. unifasciatus (Floreana)
B. snodgrassi (San Cristobal)
B. amastroides (San Cristobal)
B. exchariferus (San Cristobal)
B. nux (San Cristobal)
B. nux (Floreana)
B. sondgrassi (Española)
B. sp. 1 (Alcedo, Isabela)
100
60
83
100
99
100
100
100
66
94
100
100
95
100
100
99
100
96
79
62
0.02
100
(0.065527)
(0.209305)
(c)
Geospiza
fuliginosa
Geospiza
fortis
Geospiza
magnirostris
Geospiza
difficilis
Geospiza
scandens
Geospiza
conirostris
Camarhynchus
parvulus
Camarhynchus
pauper
Camarhynchus
psittaccula
Cactospiza
pallida
Cactospiza
heliobates
Pinaroloxias
inornata
Platyspiza
crasstrostris
Geospiza
difficilis
Certhidea
fusca
Certhidea
olivacea
0.006
100
100 100
100
100
92
93
100
77
79
67
94
76
90
65
100
100
98
96
ground finches
tree finches
Cocos
finch
Vege-
tarian
sharp
beaked
warbler finches
(d)
B. Pinzonensis (Pinzon)
undescribed
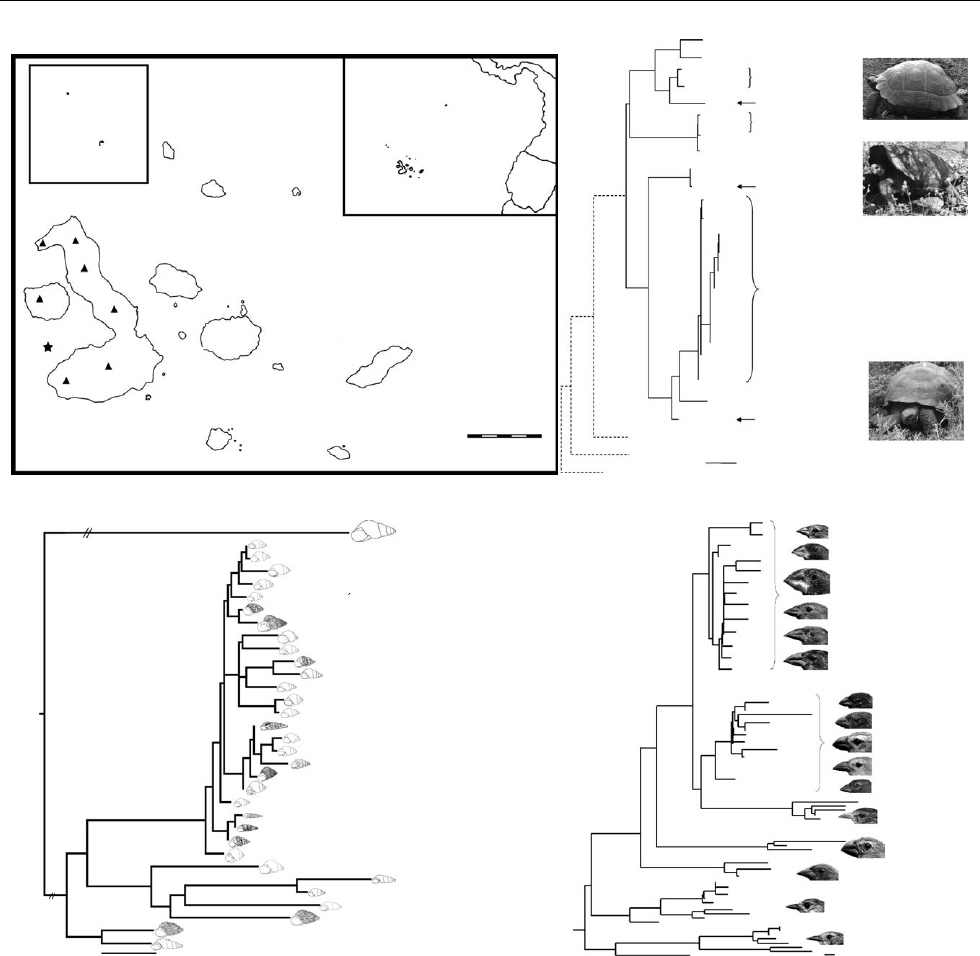
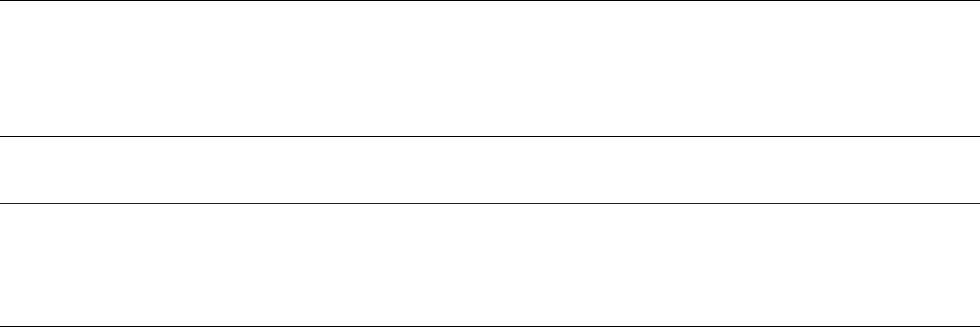
Figure 1. (a) Map of the Gala
´
pagos Islands and current hypotheses for the phylogenetic relationships of three Gala
´
pagos lineages
as follows: (b) giant Gala
´
pagos tortoises, (c) terrestrial snails and (d ) Darwin’s finches. (a) Estimated minimum and maximum
geological age for each island are in parentheses (D. Geist 2005–2008, unpublished data). Filled triangles indicate the summits
of the six Isabela volcanoes. The star indicates the location of the hot spot, currently between Fernandina and volcano Cerro
Azul of Isabela (Hooft et al. 2003). (b) Bayesian tree based on mtDNA control region sequences of extant and extinct Gala
´
pagos
tortoises. Extinct taxa are asterisked. Placement of the three lineages on Santa Cruz is indicated with an arrow, and the pictures
exemplify their different carapace morphologies. Redrawn from Russello et al. (2005).(c) Best maximum likelihood
phylogenetic tree based on combined mtDNA COI and nDNA ITS1 sequence data. Two long branches were shortened and
their actual lengths are reported in brackets below them. The snail outlines are roughly proportional to actual size. Species on
older islands connect at deeper nodes, with a significant negative relationship between the depth of the clade encompassing an
islands’ species and the island’s age. For each tree, numbers above branches are Bayesian posterior probabilities. Maximum
likelihood bootstrap proportions more than 50% are given below branches for b.(d ) Phylogenetic tree of Darwin’s finches
estimated from cytochrome b sequences and neighbour joining (Petren et al. 2005). There is significant genetic structure and
paraphyly among populations of sharp-beaked ground finches and warbler finches, but lack of resolution among the
phenotypically distinct species of ground finches and tree finches. Photographs are proportional to actual size.
Review. Diversification of Gala
´
pagos terrestrial fauna C. E. Parent et al. 3349
Phil. Trans. R. Soc. B (2008)
snails is the most spectacular in the fauna in terms of
species number (Chambers 1991; Parent & Crespi
2006). None of the other nine land snail genera that
have colonized the islands have diversified into more
than four species (Smith 1966) and preliminary
phylogenetic evidence suggests that the four species of
the genus Succinea are descendants of at least two indep-
endent colonizations (B. S. Holland 2008, personal
communication). Ta b le 2 summarizes the within-
archipelago diversification of Gala
´
pagos terrestrial
fauna, and in the following sections we will describe
the main diversification patterns.
3. ORIGINS AND AGE OF GALA
´
PAGOS FAUNA
(a) Origin of the fauna
The terrestrial Gala
´
pagos fauna that has diversified on
the islands generally has close relatives on the South
American continent. This is the case for Gala
´
pagos
tortoises, which are closely related to the smaller
bodied Geochelone chilensis, or Chaco tortoise, from
South America (Caccone et al. 1999). The diversity of
Gala
´
pagos lava lizards (Microlophus spp.) is the result of
two independent colonizations, both from the west
coast of South America (Wright 1983; Kizirian et al.
2004; Benavides et al. 2007). Likewise, Wright (1983)
suggested that the leaf-toed gecko (Phyllodactylus)
diversity stems from more than one colonization from
South America; however, this suggestion has not been
tested with phylogenetic analyses including possible
mainland relatives.
The two land bird lineages that have diversified on
Gala
´
pagos do not necessarily bear a strong relationship
to relatives on the adjacent Ecuadorian mainland:
Darwin’s finches appear to be most closely related to
grassquits (Tiaris) and their allies, a group with a wide
distribution throughout Central and South America that
has undergone a similar radiation in the Caribbean (Sato
et al. 2001; Burns et al. 2002). The closest living relatives
of Gala
´
pagos mockingbirds appear to be those currently
found in North America, northern South America and
the Caribbean, rather than the geographically nearest
species in continental Ecuador (Arbogast et al. 2006).
The endemic subspecies of the yellow warbler (Dendroica
petechia aureola) probably migrated from Latin America,
not North America, but it shows little evidence of
divergence within Gala
´
pagos (Browne et al. submitted).
Several other taxa in Gala
´
pagos (e.g. flamingos,
isopods, snakes, moths) show biogeographical affinities
with populations in the Caribbean ( Thornton 1971;
Grehan 2001); but these proposed affinities have not
been tested with molecular phylogenetic analyses.
The geographical origin of very few terrestrial
invertebrate groups has been inferred with certainty.
This gap in our knowledge is mostly because few
detailed data exist on the continental distributions of
possible sister species of Gala
´
pagos endemics. Schmitz
et al. (2007) suggested that the source of the Gala
´
pagos
microlepidopteran genus Galagete is probably coastal
South America, but this remains to be tested. Likewise,
based on morphological affinities, Gala
´
pagos bulimulid
snails seem to be most closely related to continental
South American bulimulids (Breure 1979). One must
be cautious though, as the South American fauna is
rather poorly known, especially the invertebrate
fauna, and it is possible that sister clades of Gala
´
pagos
lineages have recently become extinct, undergone range
shifts or simply have yet to be identified. A striking
example of this lack of knowledge is the genus offlightless
beetles Stomion, whose non-Gala
´
pagos closest relative is
unknown (Finston & Peck 2004).
(b) Age of the fauna
Colonization events have occurred over the last 3–4 Ma
during the existence of the present islands, and might
have happened earlier when now sunken islands were
above sea level. The presence of several drowned
seamounts on the Carnegie Ridge east of the Gala
´
pagos
(Christie et al. 1992) suggests that earlier volcanic
islands may have served as stepping stones for
colonization by some of the terrestrial fauna. Because
divergence times estimated directly from molecular data
require independent calibration, all current dating
methods require that specific assumptions be made
before inferring any divergence time. These assump-
tions typically are related either to rates of molecular
evolution (molecular clock hypothesis, local clocks
models) or both rates and times (penalized likelihood,
Bayesian methods). Among the few Gala
´
pagos taxa for
which such estimations have been attempted, the
divergence time between the mainland ancestor and
island relatives seems in some cases to be older than the
age of the existing emerged islands. The divergence
between G. chilensis and the Gala
´
pagos tortoise lineage
probably occurred 6–12 Ma, before the origin of the
oldest extant Gala
´
pagos island (Caccone et al. 1999).
However, the closest extant relative does not necessarily
Table 1. Species diversity of Gala
´
pagos terrestrial animals. (For each group, the numbers of native species in the following
categories are given: undifferentiated from non-Gala
´
pagos relatives (indigenous): differentiated from a non-Gala
´
pagos relative
but have not diversified within Gala
´
pagos (single-species endemic); diversified within Gala
´
pagos (multiple-species endemics
with number of independent within-Gala
´
pagos lineages in parentheses); and total recognized native species. Numbers are not
available for all terrestrial invertebrate groups; only data for beetles and land snails are reported.)
taxonomic group indigenous single-species endemic
multiple-species endemic
(number of lineages) total
mammals 1 1 10 (1) 12
land birds 5 7 18 (2) 30
reptiles 2 3 36 (5) 41
beetles 108 108 164 (42) 380
land snails 0 5 82 (5) 87
3350 C. E. Parent et al. Review. Diversification of Gala
´
pagos terrestrial fauna
Phil. Trans. R. Soc. B (2008)

Table 2. Summary of diversification of the terrestrial fauna of Gala
´
pagos, with the number of species per lineage that evolved on the archipelago, number of colonizing species from which they
evolved, their probable geographical origin, as well as molecular estimates (millions of years) of the initial divergence within the lineage and the temporal window of divergence from their
ancestor, whether a lineage follows the progression rule pattern (see text), whether multiple species are found within an island or volcano or whether there is evidence that the lineage is an
adaptive radiation. (For invertebrates, only lineages with 10 species or more are listed.)
class taxa
number of
species or
subspecies
phylogeny
available?
number of
colonization
events
geographical
origin
time of initial
divergence
within
Gala
´
pagos
(Ma)
divergence
time with
closest extant
lineage (Ma)
progression
rule?
multiple
species per
island or
volcano?
adaptive
radiation? source
Mammalia rice rats
(Oryzomys,
Nesoryzomys,
Megaoryzomys)
at least 8 no 3? North, Central,
and South
America
? ? ? on Santa Cruz,
Isabela and
Fernandina
no Clark (1984)
Reptilia Gala
´
pagos
giant tortoise
(Geochelone nigra)
15 (11
extant)
yes 1 South America 1.5–2.0 6.0–12.0 yes on Santa Cruz
and Isabela
may be Caccone et al.
(1999, 2002),
Beheregaray
et al. (2004),
Russello et al.
(2005) and
Ciofi et al. (2006)
lava lizards
(Microlophus)
9 yes 2 South America 1.4
#
10.2
*
yes no, one
island and
one species
no
*
Wright (1983),
Lopez et al. (1992)
2.0–2.7
#,a #
Benavides et al.
(2007)3.6–4.0
#,b
snakes (Philodryas,
Alsophis)
3 no 3? South America
and Caribbean
? ? ? yes ? Thomas (1997) and
Grehan (2001)
leaf-toed geckos
(Phyllodactylus)
6 yes 3? South America ? 8.9 yes on San
Cristo
´
bal
only
no Wright (1983)
marine
(Amblyrhynchus
cristatus)and
land (Conolophus
spp.) iguanas
1 (marine)
2 (land)
yes 1? South America O10 !2 yes land iguana:
on V. Wolf
(Isabela)
may be Wyles & Sarich
(1983),
Rassmann (1997),
S. Glaberman et al.
2005–2008
(unpublished data),
G. Gentile, et al.
2005–2008
(unpublished data)
and S. Steinfartz
et al. 2005–2008
(unpublished data)
(Continued.)
Review. Diversification of Gala
´
pagos terrestrial fauna C. E. Parent et al. 3351
Phil. Trans. R. Soc. B (2008)

Table 2. (Continued.)
class taxa
number of
species or
subspecies
phylogeny
available?
number of
colonization
events
geographical
origin
time of initial
divergence
within
Gala
´
pagos
(Ma)
divergence
time with
closest extant
lineage (Ma)
progression
rule?
multiple
species per
island or
volcano?
adaptive
radiation? source
Aves Darwin’s finches
(Geospiza,
Camarhynchus,
Cactospiza,
Platyspiza,
Certhidea)
14 yes 1 South America/
Caribbean
1.6 2.0–2.3 no yes yes Sato et al. (2001),
Burns et al. (2002)
and Petren et al.
(2005)
mockingbirds
(Nesomimus)
4 yes 1 North America/
Caribbean
? 0.6–5.5 yes no no Arbogast et al. (2006)
Insecta Galagete
microlepidoptera
12 yes 1 South America 2.9–3.7 ? no yes no Schmitz et al. (2007)
Galapaganus
weevils
10 yes 1 South America 10.7–12.1 7.2 no yes no Sequeira et al. (2000)
Stomion darklings 12 yes 1 ? ? ? ? yes no Finston & Peck
(1997, 2004)
Ammophorus 12 no 2? South America ? ? ? yes ? Peck (2006)
Philatis 20 no 1 South America? ? ? ? yes ? Peck (2001)
Scaphytopius 17 no 2? ? ? ? ? yes ? Peck (2001)
Oliarus 12 no 1 ? ? ? ? yes ? Peck (2001)
Pterostichus 12 no 2? South America? ? ? ? Desender et al. (1992)
and Peck (2006)
Dagbertus ? on Santa
Cruz,
San-
tiago
and
Isabela
5? ? ? ?Peck (2001)
Blapstinus ? yes 3? South America? ? ? ? Peck (2006)
Gastropoda Bulimulus
land snails
? yes 1 South America ? ? yes Breure (1979) and
Parent & Crespi
(2006)
yes yes
a
Microlophus ‘Eastern radiation’.
b
Microlophus ‘Western Radiation’.
3352 C. E. Parent et al. Review. Diversification of Gala
´
pagos terrestrial fauna
Phil. Trans. R. Soc. B (2008)

represent the direct ancestor, which could have been,
and perhaps was, a now extinct lineage of mainland
giant tortoises. Thus, the date of the oldest divergence
within the Gala
´
pagos lineage probably provides a
more accurate estimate of the minimum time since
colonization. The oldest node within the Gala
´
pagos
tortoise radiation is a little more recent than the oldest
islands and the molecular divergence time for the
southern Isabela clade is less than 500 000 years,
which agrees with estimates of the island’s age
( Naumann & Geist 2000; Caccone et al. 2002;
Beheregaray et al. 2003b, 2004).
Data from cytochrome b mitochondrial DNA
(mtDNA) sequences indicate a time for the divergence
of the land and marine iguana lineages of less than
10 Ma (Rassmann et al. 1997), which precedes the
origin of the emerged islands. This suggests that they
first colonized the now submerged islands and then
colonized the current islands as they emerged. Genetic
work on both land and marine iguanas is revisiting
this dating (S. Glaberman, D. Lanterbecq, S. Steinfartz,
C. Marquez, K. Dion, T. Hanley, A. Caccone 2005–
2008, unpublished data; G. Gentile, M. Milinkovitch,
A. Fabiani, C. Marquez, H. L. Snell, H. M. Snell &
W. Tapia 2005–2008, unpublished data), which was
basedonanoverallmtDNArateof2%Myr
K1
extrapolated from bovid data.
Colonization of the Gala
´
pagos by ancestors of
Darwin’s finches has been estimated at 1.6–2.3 Ma
(Sato et al. 2001), well within the age of the
present islands.
Galapaganus beetles are estimated to have diverged
from their closest living, mainland relative over 7 Ma,
based on both an intrinsic (Sequeira et al. 2008)and
extrinsic clock (Sequeira et al. 2000). However, most
DNA derived estimates of the timing of interisland
colonization and within-island diversification are
consistent with the geological formation of the islands
(Sequeira et al. 2008). The earliest divergence time
within the endemic Gala
´
pagos lepidopteran Galagete
radiation was estimated at 3.3 Ma, within the time that
the present islands were available for colonization
(Schmitz et al. 2007).
For other Gala
´
pagos lineages, it is not possible to
estimate the time of divergence from their closest
relatives because there is no reliable molecular clock
for the taxonomic group and/or genes in question
(Parent & Crespi 2006), or the relevant continental
biota is so poorly known that their close relatives cannot
be identified (table 2). Another possibility is that the
closest extant relative outside the archipelago does not
necessarily represent the direct ancestor of the insular
lineage, which could be extinct (Emerson 2002).
4. TIMING AND SEQUENCE OF COLONIZATION
AND SPECIATION
The sequence of colonization and speciation within
the archipelago can be inferred by comparing phyloge-
netic information on a lineage with the geological ages of
the islands. The most commonly predicted and tested
colonization scenario is a movement from old to young
islands, suggesting that successful intraarchipelago
colonization is more likely to occur on recently formed
islands with unoccupied niches (Brooks & McLennan
1991). This pattern, often referred to as the progression
rule (Wagner & Funk 1995), has been observed in a range
of lineages and island systems (Fleischer et al. 1998; Juan
et al. 2000). On Gala
´
pagos, this implies a diversification
sequence from southeast to northwest, paralleling the
geological formation of the islands. The Gala
´
pagos
archipelago is formed by geographical clusters of islands
of similar age (White et al. 1993)andthereforeifthe
progression rule applies, species formation should also
occur in ‘clusters’ and not necessarily match perfectly the
sequence from old to young islands.
The historical biogeographical scenarios proposed for
Gala
´
pagos endemic organisms based on phylogenetic
hypotheses suggest inconsistent relationships between
population diversification and island formation across
lineages. The Gala
´
pagos giant tortoises lineage is among
the best-studied cases of diversification on the islands,
with species-level phylogenetic analyses based on
multiple mtDNA markers (Caccone et al. 1999, 2002),
as well as population level demographic history and
population structure analyses based on both mtDNA
(Beheregaray et al. 2004; Russello et al. 2007a)and
several microsatellite markers (Beheregaray et al.
2003a,b; Ciofi et al. 2006). Although the lineage of
giant tortoises does not follow a strict linear migration
from older to younger islands, the species-level phyloge-
netic pattern uncovered from mtDNA data suggests that
giant tortoises diversified approximately in parallel to
island geological formation (Caccone et al. 2002;
figure 1b). Furthermore, estimation of population-level
genetic diversity, population divergence and recent
demographic history of Gala
´
pagos giant tortoises based
on mtDNA revealed a pattern of lineage sorting
consistent with the temporal formation of the archipelago
(Beheregaray et al. 2004). Indeed, tortoise species from
older islands have exclusively endemic haplotypes that
define divergent monophyletic clades, whereas taxa
from younger islands (Isabela, in particular) have
haplotypes shared among lineages on different volcanoes
and which exhibit a recent history of coalescence
(Beheregaray et al. 2004). One of the two lineages of
Gala
´
pagos lava lizards has also diversified in concordance
with the geological formation of island clusters of similar
age (Kizirian et al. 2004).
In a population genetic study of 22 populations of
marine iguanas (Amblyrhynchus cristatus), Rassmann
et al. (1997) found that the mtDNA cytochrome b
marker had a signature of population differentiation
concordant with geographical isolation of populations
across the archipelago. However, data from three
nuclear microsatellite loci indicated relatively high
levels of gene exchange among populations (mean
F
ST
Z0.1), contradicting the mtDNA pattern. This
discrepancy was suggested as being due to sex-biased
dispersal, with higher interpopulation dispersal in
males than females (Rassmann et al. 1997). A current
analysis of the population structure of marine iguanas
based on 13 microsatellite loci, however, depicts a very
different picture in which most island populations
represent different genetic clusters (Steinfartz et al.
2007; S. Steinfartz, D. Lanterbecq, C. Marquez &
A. Caccone 2005–2008, unpublished data). A compa-
nion analysis using mtDNA control region data reveals
Review. Diversification of Gala
´
pagos terrestrial fauna C. E. Parent et al. 3353
Phil. Trans. R. Soc. B (2008)

three main mtDNA lineages, as follows: one on the
eastern and central islands, one restricted to the northern
and western islands and one that spans from east
to west (S. Glaberman, D. Lanterbecq, S. Steinfartz,
C. Marquez, K. Dion, T. Hanley & A. Caccone
2005–2008, unpublished data). The evolutionary
relationships among these groups are unclear owing to
shallow levels of divergence, as in the cytochrome b data.
Thus, the patterns of island colonization are not as well
defined as for the tortoises. It is clear, however, that the
lineage occupying the northern and western regions of the
archipelago is the product of a recent expansion based on a
unimodal distribution of pairwise differences
between individuals as well as a high abundance of low-
frequency haplotypes. This conclusion fits with the
overall younger age of the western islands.
Gala
´
pagos mocking birds were inferred to follow the
progression rule (Arbogast et al. 2006). By contrast,
Darwin’s finches show limited evidence of diversifica-
tion closely associated with the geological formation of
the islands. Grant & Grant (2008) suggested that the
radiation of finches unfolded with an increase of types
of species with change in climate and increased habitat
diversity, associated with an increased number of
islands. However, there is no clear evidence suggesting
that the number of islands has increased through
time. Nevertheless, divergence time within this lineage
is not associated with the age of the islands where the
species are found, perhaps a result of considerable
interisland dispersal.
A progression rule pattern has been inferred for
Gala
´
pagos bulimulid land snails (Parent & Crespi 2006;
figure 1c). By contrast, Gala
´
pagos flightless weevils
(genus Galapaganus) do not follow the progression rule.
Phylogenies based on mtDNA (Sequeira et al. 2008)or
mtDNA combined with nDNA (Sequeira et al. 2008)
indicate that colonization and diversification in this
lineage are linked to geological history in a more complex
scenario. For example, Galapaganus williamsi,which
occurs only on Isabela, coalesces at the deepest node of
the phylogeny, while species from the oldest islands of
Espan
˜
ola and San Cristobal are shallower in the
phylogenetic tree. The Microlepidoptera genus Galagete
(12 species) is another example in which the progression
rule fails to explain phylogeographical patterns of
diversification. Species connecting at the deepest node
are not found on the oldest islands and the sequence of
colonization and speciation does not follow the geological
formation of the islands, but rather a stochastic
colonization scenario (Schmitz et al. 2007).
In general, the diversification sequence in Gala
´
pagos
terrestrial faunal groups seems to follow the pro-
gression rule, with the more vagile lineages constituting
the exception (Gala
´
pagos finches, Galagete lepidopter-
ans and Galapaganus weevils).
5. THE GEOGRAPHICAL MODE OF SPECIATION
In an insular context, speciation can happen between or
within islands. If colonization rate between islands is
high and individuals interbreed among island popu-
lations, these populations will effectively be part of
the same gene pool. However, ongoing but low levels
of gene flow do not constrain local adaptations of
Darwin’s finch (Petren et al. 2005) or lava lizard
(Jordan et al. 2005) populations on islets peripheral to a
main island. Lineages that have recently colonized the
islands might not have had the time to differentiate
within the archipelago. The Gala
´
pagos hawk is thought
to have diverged from its continental sister clade less
than 300 000 years ago, and populations from different
islands are genetically very similar (Bollmer et al.
2006). However, mitochondrial and minisatellite data
indicate a general pattern of rapid population expan-
sion followed by genetic isolation of hawk populations,
suggesting that this lineage is in the earliest stages of
further divergence (Bollmer et al. 2006). Genetic
analysis of the Gala
´
pagos hawks’s ischnoceran louse
may shed further light on the cryptic evolutionary
history of its host (Whiteman et al. 2007).
(a) Speciation between islands
If colonization rate between islands is low, one would
expect to find a pattern of genetic structuring among
island populations, potentially leading to differentiated
species that are single island endemics. Rassmann
et al. (1997) initially proposed that marine iguana
populations across the archipelago showed very little
divergence but ongoing work suggests that most
island populations have considerable levels of
genetic differentiation (S. Steinfartz, D. Lanterbecq,
C. Marquez & A. Caccone 2005–2008, unpublished
data). Gala
´
pagos giant tortoises have diverged to form
15 taxa, 11 of which are extant. Their taxonomic
status is still in flux. Some authors consider them
different subspecies (Pritchard 1996), while others
refer to them as species owing to their genetic
distinctiveness (Powell & Caccone 2006; Russello
et al. 2007a). Espan
˜
ola, San Cristobal, Pinzo
´
n,
Santiago and Pinta each has only one named species;
San Cristobal had two distinct populations but one is
now extinct (Caccone et al. 2002; Russello et al.
2007a). Isolation among islands has promoted specia-
tion in the two independent lineages of lava lizards
(Kizirian et al. 2004; Benavides et al. 2007). Using
microsatellite markers, Jordan & Snell (2008) have
tested the hypothesis that genetic drift in isolation has
caused the differentiation of Gala
´
pagos lava lizards on
12 islets that were probably connected to the larger
island of Santa Cruz during the Late Pleistocene.
They found strong genetic differentiation among
island populations, a positive correlation of genetic
diversity with island size, no relationship between
genetic and geographical distance and a strong
negative correlation between heterozygosity and
measures of genetic differentiation. They concluded
that overwater dispersal is rare and further differen-
tiation of lava lizards may be prevented by island
fusion during periods of lower sea level. The land
iguanas (Snell et al. 1984; Rassmann 1997) and the
mocking birds (Arbogast et al. 2006) also have at
most one species per island, but the relative contri-
butions ofselection and drift to differentiation remain
unclear. Recent work on the two Gala
´
pagos land
iguanas, Conolophus subcristatus and Conolophus
pallidus (G. Gentile, M. Milinkovitch, A. Fabiani,
C. Marquez, H. L. Snell, H. M. Snell & W. Tapia
2008–2008, unpublished data), based on mtDNA
3354 C. E. Parent et al. Review. Diversification of Gala
´
pagos terrestrial fauna
Phil. Trans. R. Soc. B (2008)

sequence data, confirms the genetic discontinuities
between the populations of C. subcristatus from the
western (Fernandina and Isabela) and central (Santa
Cruz, Baltra, Seymour and Plaza Sur) islands. This
pattern seems to parallel that of their sister taxon, the
marine iguanas, in which there is clear genetic
differentiation between eastern and western islands
(S.Glaberman,D.Lanterbecq,S.Steinfartz,
C.Marquez,K.Dion,T.Hanley&A.Caccone
2005–2008, unpublished data; S. Steinfartz, D. Lanter-
becq, C. Marquez & A. Caccone 2005–2008, unpub-
lished data). The level of differentiation between the two
C. subcristatus groups is only slightly lower than that
between the two species (C. subcristatus and C. pallidus).
Darwin’s finches show limited evidence of speciation
on different islands. The larger central islands contain
largely the same suite of 9–10 species, many of which
also occur on several smaller and peripheral islands
(Grant 1999). Several phylogenetic analyses have
revealed little resolution among six species of tree
finches and five species of ground finches (Freeland &
Boag 1999; Sato et al. 2001; Petren et al. 2005;
figure 1d ). A combination of factors is probably
responsible for this, including the recent origin of
many species, the homogenizing effects of hybrid-
ization (Grant et al. 2004) and interisland dispersal
(Petren et al. 2005). Allopatric speciation has been
inferred between the small (Geospiza scandens)and
large (Geospiza conirostris) cactus finches, although
both populations of large cactus finches are more
closely related genetically to different populations of
small cactus finches than they are to one another.
The warbler finches (Certhidea olivacea and Certhidea
fusca) were believed to be the same species, until
studies showed different subsets of islands to harbour
two different species that are more genetically distinct
than any other species in the entire group (Petren
et al. 1999; Tonnis et al. 2005). There is pronounced
morphological divergence between-island populations
of several species of Darwin’s finches. Some of these
differences are paralleled by substantial genetic
differences (Geospiza difficilis, G. scandens/G. conirostris),
while in other cases, genetic differences are minimal or
are not correlated with morphological or geographical
divergence (Geospiza fortis/Geospiza magnirostris; Petren
et al. 1999, 2005). Recent studies suggest interisland
movements of Darwin’s finches may not be as rare as
previously suspected (Petren et al. 2005; Tonnis et al.
2005). Morphological differences among island popu-
lations may therefore not be as much evidence of isolation
from gene flow as previously thought, but rather evidence
of differing selective regimes and local adaptation in the
presence of gene flow. Theoretically, low levels of gene
exchange may promote local adaptation under
some conditions (Whitlock & Phillips 2000). The
warbler finches (C. fusca and C. olivacea)appearto
conform to an allopatric mode of speciation, being clearly
separated on different subsets of islands. However, the
occurrence of some very widely distributed C. fusca
haplotypes suggests that speciation was not driven by
geographical isolation and limited dispersal. Other
factors, such as habitat or natural selection against
immigrants, may have limited genetic exchange and
allowed speciation to occur (To nn i s et al. 2005).
Terrestrial invertebrates often have low dispersal
ability (especially flightless groups), so single island
endemics would be expected. The flightless beetle
genus Neoryctes includes four single island endemic
species, each restricted to the moist highland zone
of the island on which it occurs (Cook et al. 1995). The
microlepidopteran genus Galagete has a complex pattern
of diversification. Some of the species are single-island
endemics, others are found on multiple islands, and two
occur on 10 and 11 islands (Schmitz et al. 2007). The
inference of the geographical mode of speciation from
the phylogeny is problematic, mainly owing to the
wide distribution of most of its taxa. Little is known
of the ecology of Galagete (Schmitz et al. 2007), which
makes it difficult to infer ecological or habitat shifts.
In sum, patterns of speciation between islands tend
largely to reflect a combination of dispersal capability
and habitat suitability. Less vagile taxa show more
pronounced genetic structure among island popu-
lations. They are more prone to longer periods of
absence from suitable islands as a result of extinction.
More vagile taxa show little differentiation among
subsets of islands, but may be absent from islands with
apparent suitable habitat. Several taxa show non-
overlapping distributions among subsets of islands.
In these cases, co-occurrence may be limited by
dispersal, but also by the presence of other similar
species. The possibility of occasional gene exchange
between islands that would create loose evolutionary
connections among differentiated populations remains
largely unexplored.
(b) Speciation within islands
In addition to island isolation, other physical features
of islands can influence the mode of diversification in
an archipelago. Island area and elevation are positively
correlated in many island groups and are prime
determinants of moisture and island habitat diversity.
Greater habitat diversity provides the potential for
within-island diversification, but also increases the
chances that a new colonist will find a suitable habitat.
Once a particular lineage has colonized an island, it can
speciate within that island either by adapting to
different habitats, commonly referred to as ecological
speciation (Schluter 2000), or by allopatric speciation if
an island is large enough for populations to become
isolated by geographical or topological means (volca-
noes, barren lava, inhospitable habitat, etc.). The
strength of the effect of island area depends not only
on geography and topology but also on the dispersal
ability of the organisms, being strongest for animals
with low dispersal ability and specialized ecological
requirements. The low dispersal ability of the giant
tortoises is a good example, with four species on
Isabela, one on each major volcano. Isabela probably
started as separate island volcanoes that then
merged. The volcanoes are now separated by lava
fields, which are a formidable obstacle to tortoise
movement. It is therefore possible that their popu-
lations were isolated for 400 000 years, and then
experienced limited mixing (over the lava fields) for
the past 100 000 years once the volcanoes were
connected. mtDNA and nDNA data support the
genetic distinctiveness of the three species living on
Review. Diversification of Gala
´
pagos terrestrial fauna C. E. Parent et al. 3355
Phil. Trans. R. Soc. B (2008)

the central and northern volcanoes (Alcedo, Darwin,
Wolf; Caccone et al. 2002; Beheregaray et al. 2004;
Ciofi et al. 2006, in press; Russello et al. 2007a).
However, genetic divergence and similarities among
populations on the southern volcanoes (Cerro Azul,
Sierra Negra) do not support the taxonomic subdivi-
sions (Ciofi et al. 2006) that were previously suggested
based on morphology ( Fritts 1984; Pritchard 1996).
The taxonomic assignment of tortoise populations on
Santa Cruz has been contentious. There are at least
three lineages that are as genetically distinct from
each other as from lineages on other islands, being
connected only through the deepest nodes in the
molecularly based tree (Russello et al. 2005). Shell
shape (domed versus saddlebacked carapace) is the
most distinctive phenotypic difference among giant
tortoise populations and has been associated with
habitat differences. The ancestral state of this character
in Gala
´
pagos is probably domed (Caccone et al. 2002;
Russello et al. 2005) and recent DNA data from extinct
taxa suggest that the saddleback morphology arose
independently more than once (Russello et al. 2005;
Poulakakis et al. 2005–2008, unpublished data).
However, the role of selection and developmental
constraints in the diversification of Gala
´
pagos giant
tortoises remains to be clearly determined. Research
using novel field friendly three-dimensional imaging
techniques and morphometric methods is addressing
such issues (Chiari et al. in press).
Within-island divergence has been suggested as a
mode of speciation for Darwin’s finches. Divergence
occurs along an elevational gradient in small ground
finches on Santa Cruz (Kleindorfer et al. 2006).
Geospiza conirostris on Genovesa showed an ephemeral
tendency to mate assortatively according to similarities
in beak shape and song (Grant & Grant 1989).
Assortative mating according to beak size occurs within
populations of medium ground finches on Santa Cruz,
although this tendency appears to be significantly
reduced by human activity (Hendry et al. 2006) and
wet conditions, when competition is also presumably
reduced (Huber et al. 2007). Sympatric speciation has
yet to be convincingly demonstrated in birds on islands
such as the Gala
´
pagos that are smaller than 10 000 km
2
(Coyne & Price 2000), and so it seems unlikely that any
of these situations will result in complete speciation.
Lack (1947) proposed a model for within-island
species build-up of Darwin’s finches that relied on
limited dispersal between islands to create initial diff-
erences in allopatry, followed by rare colonization,
establishment of secondary contact and further differ-
entiation in sympatry as a result of resource compe-
tition and character displacement. This model has in
principle been adapted to account for faunal commu-
nity build-up in a wide variety of systems including
Gala
´
pagos taxa. Despite the apparent lack of isolation
between finch populations on different islands (Petren
et al. 2005), Lack’s model of faunal build-up may
still apply. Immigrants that normally interbreed with
residents may on rare occasions form distinct, repro-
ductively isolated populations, depending on environ-
mental circumstances and the combination of traits
possessed by immigrants and residents (e.g. Grant &
Grant 2008).
In the beetle genus Galapaganus, diversification was
within island on San Cristobal, one of the oldest
islands, and within-island speciation could not be
rejected for the Santa Cruz clade; in both cases, shifts
to highland habitats seem to have evolved indepen-
dently (Sequeira et al. 2008). Although this habitat shift
is consistent with selection contributing to within-
island diversification, it is not known whether it is
associated with a corresponding adaptive shift in
phenotype. Notably, Sequeira et al. (2008) inferred
that speciation did not occur within the younger island
of Isabela, where diversity is the result of between-island
colonization. Similarly, Parent & Crespi (2006) found a
combination of within- and between-island speciation in
Gala
´
pagos bulimulid snails. Polyphyletic land snail
assemblages were found on islands centrally located in
space and time (Isabela, Santa Cruz, Pinzo
´
n, Santiago),
whereas the more isolated islands of Espan
˜
ola, San
Cristobal and Floreana (again in both space and time)
were occupied mostly by monophyletic assemblages
resulting from within-island diversification. Some buli-
mulid species occur in sympatry and may have adapted to
the different habitats where they occur (Coppois &
Glowacki 1983; C. E. Parent & B. J. Crespi 2000–2005,
unpublished data). However, the relative importance
of drift and selection in within-island diversification of
this group is not known.
The beetle genus Stomion has also diversified both
within and between islands ( Finston & Peck 2004).
Each population contains only one morphospecies,
and even on large islands such as Santa Cruz and
Isabela where two or three species occur, they never
occur in sympatry. Finston & Peck (2004) therefore
suggested that speciation happened in allopatry.
Stomion species have low vagility, even at very small
local scales ( Finston et al. 1997), which should
promote micro-allopatric speciation, in agreement
with the scenario proposed by Finston & Peck
(2004). No evidence of ecological or morphological
adaptation has been found in the genus (Finston &
Peck 1997, 2004), strengthening the idea that diversi-
fication happens via micro-allopatric speciation within
islands following between-island colonization. Peck
(1990) has suggested that troglobites (cave-dwelling
invertebrates) might provide good examples of diversi-
fication within islands. There are several examples of a
cave-dwelling form that has a sister species living
nearby outside the cave (Peck 1990; Peck & Finston
1993), but without a molecular phylogeny we cannot
tell how these species were formed.
In sum, the clearest cases of within-island speciation
occur in taxa with low vagility and on large islands with
diverse habitats. Isabela, with several volcanoes separ-
ated by recent lava flows, is especially conducive to
within-island diversification. In some cases, subsets of
taxa have found their way to other islands, making it
more difficult to distinguish within-island speciation
from multiple between-island speciation events. For
more vagile taxa, speciation between islands seems
more likely, given the extreme habitat differences
among islands; but within-island processes are often
difficult to rule out, and in either case, ecological
specialization can play an important role.
3356 C. E. Parent et al. Review. Diversification of Gala
´
pagos terrestrial fauna
Phil. Trans. R. Soc. B (2008)

(c) Dynamics of species diversification
The number of species on an island depends on
speciation, colonization and extinction rates. The
three processes are affected by biogeography (island
area, elevation and insularity) and ecology (habitat
diversity). However, the species diversity equilibrium is
dynamic and changes through time because islands
themselves change (Whittaker et al. 2008). Moreover,
a delay is expected between the time an island becomes
available for colonization and speciation, and the time
equilibrium is reached. For example, Parent & Crespi
(2006) found that the younger islands of Fernandina
and Isabela had fewer land snail species than expected
based on their area, elevation and insularity. Similarly,
there are only two species of flightless Galapaganus
beetles on Isabela, whereas San Cristobal, Santa Cruz
and Floreana each has three species (Sequeira et al.
2008). By contrast, there are more species of the moth
genus Galagete on Isabela and Fernandina (eight each)
than on any of the older islands (Schmitz et al. 2007).
This contrast among lineages might be a reflection of
their different dispersal capacity or the stochasticity of
interisland dispersal in some lineages.
Furthermore, the geographical configuration of the
whole archipelago is dynamic, and there have been
changes in island number, size and isolation due to sea-
level variation on various time scales. On a short time
scale (from now back to the Pleistocene), it appears
that changes in sea levels created terrestrial avenues for
gene flow during glacial periods that disappeared when
sea levels rose. The effect this sea level change can have
on species diversity on Gala
´
pagos has been relatively
unexplored, but Jordan & Snell (2008) suggested that
lava lizard diversity might have increased if islands
had been isolated for longer periods. Environmental
changes over the short term can influence the
evolutionary trajectory of species (Grant 1999), so it
is also likely that longer term climate changes have had
an important effect on the evolution and extinction of
species that remains largely unknown. A sobering
example comes from a recent study on El Nino effects
on levels of genetic diversity in Gala
´
pagos marine
iguanas (Steinfartz et al. 2007), in which even species
with large population sizes may suffer dramatic
collapses in genetic diversity when populations are
subject to frequent and possibly recurrent environ-
mentally devastating events.
6. FUTURE PROSPECTS FOR EVOLUTIONAR Y
RESEARCH ON GALA
´
PAGOS ISLANDS
Despite the large volume of evolutionary research that
has been undertaken in Gala
´
pagos, several species
groups remain relatively unstudied in terms of diversi-
fication within Gala
´
pagos and their relationship to
possible sources of colonists (table 2). Most phyloge-
netic studies of Gala
´
pagos organisms have been based
on mtDNA, sometimes in combination with one or a
few nuclear markers. Mitochondrial DNA has desir-
able properties for phylogenetic inference (Avise
2000), but the likelihood that a single locus reflects
the history of species and populations decreases when
divergence occurs rapidly and recently (Hudson &
Coyne 2002), as for much of the diversification in the
Gala
´
pagos. Multilocus studies of sequence variation,
such as intron locus variation (Edwards et al. 2007),
offers some promise for providing more robust
estimates of phylogenetic and phylogeographic diver-
gence. However, in the only case in which they have
been used, giant tortoises, they were not helpful
(Caccone et al. 2004). For more vagile taxa such as
winged insects and birds, multilocus microsatellites
now offer a powerful means to estimate even low levels
of gene flow, admixture and population divergence.
Ecology and habitat specialization can be critical
factors in speciation both within and between islands.
Phylogenetic studies can be broad but are limited in their
ability to reveal speciation processes. The number of
studies of natural history, ecology, evolution and
behaviour is small and limited in scope. Field studies
are limited spatially and temporally by logistics and
funding. Thus a great deal remains to be revealed about
speciation in Gala
´
pagos animals. One need only consider
a single field research programme (Grant 1999; Grant &
Grant 2008) to see the magnitude of contributions to
the study of evolution and speciation that are possible.
7. CONSERVATION AND THE ROLE
OF EVOLUTIONARY GENETICS IN
GALA
´
PAGOS RESEARCH
While providing ecologists and evolutionary biologists
with ‘natural laboratories’, island biotas are also very
often in danger of going extinct. This is due both to
their generally small population sizes associated with
small geographical ranges, as well as to the relatively
simple ecosystems they inhabit (which is why they
are so valued by biologists). Island ecosystems are
therefore particularly susceptible to catastrophic
disturbances by invasive species. Understanding the
phylogenetics and biogeography of island species
allows greater insights into setting conservation
priorities and enacting rational conservation policies,
as illustrated in the examples below on Gala
´
pagos
tortoises (Powell & Caccone 2008).
Phylogenetic analysis detected previously unrecog-
nized taxa that deserve special conservation attention
(Russello et al. 2005) and assigned individuals of
unknown origin (zoo animals) to their correct location
(Burns et al. 2003; Russello et al. 2007b). Such studies
also allowed identification of genetic lineages in
natural populations that were previously thought to
be extinct or nearly extinct (Russello et al. 2007a;
Poulakakis et al. 2005–2008, unpublished data).
Understanding the genetics of endangered island
biota may also aid in designing more rational recovery
efforts such as captive breeding/repatriation pro-
grammes (Milinkovitch et al. 2004, 2007).
Over the last century, the human population of
Gala
´
pagos has risen dramatically. Prior to 1900, fewer
than 300 people lived in the islands (Lundh 2001).
There are now nearly 30 000 residents on four
inhabited islands and more than 130 000 visitors
each year ( Watkins & Cruz 2007). The human
population boom has caused a marked increase in
introduced species and diseases (Fessl & Tebbich
2002; Wikelski et al. 2004; Gottdenker et al. 2005).
Several populations of birds have recently become
Review. Diversification of Gala
´
pagos terrestrial fauna C. E. Parent et al. 3357
Phil. Trans. R. Soc. B (2008)

extinct (Grant 1999; Grant et al. 2005) and several
species are now threatened or on the brink of
extinction (e.g. Dvorak et al. 2004).
The near future will determine whether the
Gala
´
pagos will provide a model for the deterioration
of an island fauna in our time, or whether the brighter
prospect of effective management and persistence will
be realized so that future generations can continue to
study the processes of evolution in action.
We thank the staff of the Charles Darwin Research Station
and Parque Nacional de Gala
´
pagos for their continued
support and collaboration, and all the volunteers and
undergraduate and graduate students who have collaborated
in field and laboratory work. We are grateful to E. Benavides,
F. Breden, B. Emerson, D. Geist, G. Gentile, S. Glaberman,
A. Hendry, M. A. Jordan, L. F. Keller, J. R. Powell,
P. Schmitz, A. Sequeira, F. Sulloway and S. Steinfartz, and
an anonymous reviewer for providing comments and for
sending unpublished results. We thank Michael Russello for
providing figure 1b. We also thank R. H. Cowie and S. A.
Trewick for their helpful comments and for giving us the
opportunity to contribute this review. A.C.’s work was
supported by NSF (DEB 322672 to J. R. Powell), YIBS-
Ecosave, the National Geographic Society, the Paul and Bay
Foundation and the AWARE Foundation; C.E.P.’s work was
supported by le Fonds Nature et Technologies du Que
´
bec
(FCAR), the Natural Sciences and Engineering Research
Council of Canada, Unitas Malacologica and the Conchol-
ogists of America; K.P.’s work was supported by NSF
(DEB 0317687).
REFERENCES
Allan, R., Lindesay, J. & Parker, D. 1996 El Nin˜o—southern
oscillation and climatic variability. CSIRO Publishing:
Collingwood, Victoria, Australia.
Arbogast, B. S., Drovetski, S. V., Curry, R. L., Boag, P. T.,
Seutin, G., Grant, P. R., Grant, B. R. & Anderson, D. J.
2006 The origin and diversification of Gala
´
pagos mock-
ingbirds. Evolution 60, 370–382. (doi:10.1554/03-749.1)
Avise, J. C. 2000 Phylogeography: the history and formation of
species. Cambridge, MA: Harvard University Press.
Bailey, K. 1976 Potassium-Argon ages from the Gala
´
pagos
Islands. Science 192, 465–466. (doi:10.1126/science.192.
4238.465)
Beheregaray, L. B., Ciofi, C., Caccone, A., Gibbs, J. P. &
Powell, J. R. 2003a Genetic divergence, phylogeography
and conservation units of giant tortoises from Santa Cruz
and Pinzo
´
n, Gala
´
pagos Islands. Conserv. Genet. 4, 31–46.
(doi:10.1023/A:1021864214375)
Beheregaray, L. B., Ciofi, C., Geist, D., Gibbs, J. P., Caccone,
A. & Powell, J. R. 2003b Genes record a prehistoric
volcano eruption in the Gala
´
pagos. Science 302, 75.
(doi:10.1126/science.1087486)
Beheregaray, L. B., Gibbs, J. P., Havill, N., Fritts, T. H.,
Powell, J. R. & Caccone, A. 2004 Giant tortoises are not so
slow: rapid diversification and biogeographic consensus in
the Gala
´
pagos. Proc. Natl Acad. Sci. USA 101, 6514–6519.
(doi:10.1073/pnas.0400393101)
Benavides, E., Baum, R., McClellan, D. & Sites Jr, J. W. 2007
Molecular phylogenetics of the lizard genus Microlophus
(Squamata: Tropiduridae): aligning and retrieving indel
signal from nuclear introns. Syst. Biol. 56, 776–797.
(doi:10.1080/10635150701618527)
Bollmer, J. L., Kimball, R. T., Whiteman, N. K., Sarasola,
J. H. & Parker, P. G. 2006 Phylogeography of the
Gala
´
pagos hawk (Buteo galapagoensis): a recent arrival to
the Gala
´
pagos Islands. Mol. Phylogenet. Evol. 39, 237–247.
(doi:10.1016/j.ympev.2005.11.014)
Breure, A. S. H. 1979 Systematics, phylogeny and zoogeography
of Bulimulinae (Mollusca). Leiden, Germany: Zoologische
Verhandelingen.
Brooks, D. R. & McLennan, D. A. 1991 Phylogeny, ecology,
and behavior: a research program in comparative biology.
Chicago, IL: University of Chicago Press.
Browne, R. A., Collins, E. I. & Anderson, D. J. Submitted.
Genetic structure and morphological variation among
Gala
´
pagos populations of the yellow warbler.
Burns, K. J., Hackett, S. J. & Klein, N. K. 2002 Phylogenetic
relationships and morphological diversity in Darwin’s
finches and their relatives. Evolution 56, 1240–1252.
(doi:10.1111/j.0014-3820.2002.tb01435.x)
Burns, C. T., Ciofi, C., Beheregaray, L. B., Fritts, T. H.,
Gibbs, J. P., Marquez, C., Milinkovitch, M. C., Powell,
J. R. & Caccone, A. 2003 The origin of captive Gala
´
pagos
tortoises based on DNA analysis: implications for the
management of natural populations. Anim. Conserv. 6,
329–337. (doi:10.1017/S1367943003003408)
Caccone, A., Gibbs, J. P., Ketmaier, V., Suatoni, E. & Powell,
J. R. 1999 Origin and evolutionary relationships of giant
Gala
´
pagos tortoises. Proc. Natl Acad. Sci. USA 96,
13 223–13 228. (doi:10.1073/pnas.96.23.13223)
Caccone, A., Gentile, G., Gibbs, J. P., Fritts, T. H., Snell,
H. L., Betts, J. & Powell, J. R. 2002 Phylogeography and
history of giant Gala
´
pagos tortoises. Evolution 56,
2052–2066. (doi:10.1111/j.0014-3820.2002.tb00131.x)
Caccone, A., Gentile, G., Burns, C., Sezzi, E., Bergman, W.
& Powell, J. R. 2004 Extreme difference in rate of
mitochondrial and nuclear DNA evolution in a large
ectotherm, Gala
´
pagos tortoises. Mol. Phylogenet. Evol. 31,
794–798. (doi:10.1016/j.ympev.2004.02.004)
Chambers, S. M. 1991 Biogeography of Gala
´
pagos land
snails. In Gala
´
pagos marine invertebrates (ed. M. J. James),
pp. 307–325. New York, NY: Plenum.
Chiari, Y., Wang, B., Rushmeier, H. & Caccone, A. In press.
Using digital images to reconstruct 3D biological forms:
a new tool for morphometric studies. Biol. J. Linn. Soc.
Christie, D. M., Duncan, R. A., McBirney, A. R., Richards,
M. A., White, W. M., Harpp, K. S. & Fox, C. G. 1992
Drowned islands downstream from the Gala
´
pagos hotspot
imply extended speciation times. Nature 355, 246–248.
(doi:10.1038/355246a0)
Ciofi, C., Wilson, G. A., Beheregaray, L. B., Marquez, C.,
Gibbs, J. P., Tapia, W., Snell, H. L., Caccone, A. & Powell,
J. R. 2006 Phylogeographic history and gene flow
among giant Gala
´
pagos tortoises on southern Isabela
Island. Genetics 172, 1727–1744. (doi:10.1534/genetics.
105.047860)
Ciofi, C., Caccone, A., Beheregaray, L. B., Milinkovitch, M.
C., Russello, M. A. & Powell, J. R. In press. Genetics and
conservation on islands: the Gala
´
pagos giant tortoise as a
case study. In Population genetics for animal conservation
(eds G. Bertorelle, M. W. Bruford, C. Chemini, H. C.
Hauffe & C. Vernesi). Cambridge, UK: Cambridge
University Press.
Clark, D. A. 1984 Native land mammals. In Key environments:
Gala
´
pagos (ed. R. Perry), pp. 225–231. Oxford, UK:
Pergamon Press.
Cook, J., Howden, H. F. & Peck, S. B. 1995 The Gala
´
pagos-
Islands genus Neoryctes Arrow (Coleoptera, Scarabaeidae,
Dynastinae). Can. Entomol. 127 , 177–193.
Coppois, G. & Glowacki, C. 1983 Bulimulid land snails from
the Gala
´
pagos: 1. Factor analysis of Santa Cruz Island
species. Malacologia 23, 209–219.
Cox, A. 1983 Ages of the Gala
´
pagos Islands. In Patterns of
Gala
´
pagos organisms (eds R. I. Bownman, M. Berson &
A. E. Leviton), pp. 11–23. San Franscisco, CA: American
Association for the Advancement of Science, Pacific
Division.
3358 C. E. Parent et al. Review. Diversification of Gala
´
pagos terrestrial fauna
Phil. Trans. R. Soc. B (2008)

Coyne, J. A. & Price, T. D. 2000 Little evidence for sympatric
speciation in island birds. Evolution 54, 2166–2171.
(doi:10.1111/j.0014-3820.2000.tb01260.x)
Darwin, C. R. 1845 Journal of researches into the natural history
and geology of the countries visited during the voyage of
H.M.S. Beagle round the world, under the Command of Capt.
FitzRoy, R.N. London, UK: John Murray.
Darwin, C. R. 1859 On the origin of species by means of natural
selection; or the preservation of favoured races in the struggle for
life. London, UK: John Murray.
Desender, K., Baert, L. & Maelfait, J.-P. 1992 Distribution
and speciation of carabid beetles in the Gala
´
pagos
archipelago ( Ecuador). Bull. Inst. R. Sci. Natl Belg. 62,
57–65.
Dvorak, M., Vargas, H., Fessl, B. & Tebbich, S. 2004 On the
verge of extinction: a survey of the mangrove finch
Cactospiza heliobates and its habitat on the Gala
´
pagos
Islands. Oryx 38, 171–179. (doi:10.1017/S00306053040
00316)
Edwards, S. V., Liu, L. Y. & Pearl, D. K. 2007 High-
resolution species trees without concatenation. Proc. Natl
Acad. Sci. USA 104, 5936–5941. (doi:10.1073/pnas.
0607004104)
Emerson, B. C. 2002 Evolution on oceanic islands: molecular
phylogenetic approaches to understanding pattern and
process. Mol. Ecol. 11 , 951–966. (doi:10.1046/j.1365-
294X.2002.01507.x)
Fessl, B. & Tebbich, S. 2002 Philornis downsi - a recently
discovered parasite on the Gala
´
pagos archipelago—a
threat for Darwin’s finches? Ibis 144, 445–451. (doi:10.
1046/j.1474-919X.2002.00076.x)
Finston, T. L. & Peck, S. B. 1997 Genetic differentiation and
speciation in Stomion (Coleoptera: Tenebrionidae): flight-
less beetles of the Gala
´
pagos islands, Ecuador. Biol.
J. Linn. Soc. 61, 183–200.
Finston, T. L. & Peck, S. B. 2004 Speciation in Darwin’s
darklings: taxonomy and evolution of Stomion beetles in
the Gala
´
pagos Islands, Ecuador (Insecta: Coleoptera:
Tenebrionidae). Zool. J. Linn. Soc. 141, 135–152. (doi:10.
1111/j.1096-3642.2004.00115.x)
Finston, T. L., Peck, S. B. & Perry, R. B. 1997 Population
density and dispersal ability in Darwin’s darklings:
flightless beetles of the Gala
´
pagos Islands. Pan-Pac.
Entomol. 73, 110–121.
Fleischer, R. C., McIntosh, C. E. & Tarr, C. L. 1998
Evolution on a volcanic conveyor belt: using phylogeo-
graphic reconstructions and K–Ar-based ages of the
Hawaiian Islands to estimate molecular evolutionary
rates. Mol. Ecol. 7, 533–545. (doi:10.1046/j.1365-294x.
1998.00364.x)
Freeland, J. R. & Boag, P. T. 1999 Phylogenetics of Darwin’s
finches: paraphyly in the tree-finches, and two divergent
lineages in the warbler finch. Auk 116, 577–588.
Fritts, T. H. 1984 Evolutionary divergence of giant tortoises
in Gala
´
pagos. Biol. J. Linn. Soc. 21, 165–176. (doi:10.
1111/j.1095-8312.1984.tb02059.x)
Geist, D. 1996 On the emergence and submergence of the
Gala
´
pagos Islands. Not. Gala
´
pagos 56 , 5–9.
Gottdenker, N. L., Walsh, T., Vargas, H., Merkel, J., Jimenez,
G. U., Miller, R. E., Dailey, M. & Parker, P. G. 2005
Assessing the risks of introduced chickens and their
pathogens to native birds in the Gala
´
pagos archipelago.
Biol. Conserv. 126, 429–439. (doi:10.1016/j.biocon.2005.
06.025)
Grant, P. R. 1999 Ecology and evolution of Darwin’s finches.
Princeton, NJ: Princeton University Press.
Grant, B. R. & Grant, P. R. 1993 Evolution of Darwin’s
finches caused by a rare climatic event. Proc. R. Soc. B 251,
111–117. (doi:10.1098/rspb.1993.0016)
Grant, P. R. & Grant, B. R. 1989 Sympatric speciation in
Darwin’s finches. In Speciation and its consequences (eds
D. Otte & J. A. Endler), pp. 433–457. Sunderland, MA:
Sinauer.
Grant, P. R. & Grant, B. R. 2008 How and why species multiply:
the radiation of Darwin’s finches. Princeton, NJ: Princeton
University Press.
Grant, P. R., Grant, B. R., Markert, J. A., Keller, L. F. &
Petren, K. 2004 Convergent evolution of Darwin’s finches
caused by introgressive hybridization and selection.
Evolution 58, 1588–1599. (doi:10.1554/04-016)
Grant, P. R., Grant, B. R., Petren, K. & Keller, L. F. 2005
Extinction behind our backs: the possible fate of one of
the Darwin’s finch species on Isla Floreana, Gala
´
pagos.
Biol. Conserv. 122, 499–503. (doi:10.1016/j.biocon.2004.
09.001)
Grehan, J. 2001 Biogeography and evolution of the
Gala
´
pagos: integration of the biological and geological
evidence. Biol. J. Linn. Soc. 74, 267–287. (doi:10.1006/
bijl.2001.0576)
Hall, M. L. 1983 Origin of Espan
˜
ola Island and the age of
terrestrial life on the Gala
´
pagos Islands. Science 221,
545–547. (doi:10.1126/science.221.4610.545)
Hendry, A. P., Grant, P. R., Grant, B. R., Ford, H. A.,
Brewer, M. J. & Podos, J. 2006 Possible human impacts on
adaptive radiation: beak size bimodality in Darwin’s
finches. Proc. R. Soc. B 273, 1887–1894. (doi:10.1098/
rspb.2006.3534)
Hickman, C. & Lipps, J. H. 1985 Geologic youth of
Gala
´
pagos Islands confirmed by marine stratigraphy and
paleontology. Science 277, 1578–1580. (doi:10.1126/
science.227.4694.1578)
Hoernle, K., van den Bogaard, P., Werner, R., Lissinna, B.,
Hauff, F., Alvarado, G. & Garbe-Schonberg, D. 2002
Missing history (16–71 Ma) of the Galpapagos hotspot:
implications for the tectonic and biological evolution of
the Americas. Geology 30, 795–798. (doi:10.1130/0091-
7613(2002)030!0795:MHMOTGO2.0.CO;2)
Hooft, E. E., Toomey, D. R. & Solomon, S. C. 2003
Anomalously thin transition zone beneath the Gala
´
pagos
hotspot. Earth Planet. Sci. Lett. 216, 55–64. (doi:10.1016/
S0012-821X(03)00517-X)
Huber, S. K., De Leo
´
n, L. F., Hendry, A. P., Bermingham, E.
& Podos, J. 2007 Reproductive isolation of sympatric
morphs in a population of Darwin’s finches. Proc. R. Soc.
B 274, 1709–1714. (doi:10.1098/rspb.2007.0224)
Hudson, R. R. & Coyne, J. A. 2002 Mathematical
consequences of the genealogical species concept.
Evolution 56, 1557–1565. (doi:10.1111/j.0014-3820.
2002.tb01467.x)
Jordan, M. A. & Snell, H. L. 2008 Historical fragmentation
of islands and genetic drift in populations of Gala
´
pagos
lava lizards (Microlophus albermarlensis complex). Mol.
Ecol. 14, 859–867. (doi:10.1111/j.1365-294X.2005.
02452.x)
Jordan, M. A., Snell, H. L., Snell, H. M. & Jordan, W. C.
2005 Phenotypic divergence despite high levels of gene
flow in Gala
´
pagos lava lizards (Microlophus albemarlensis).
Mol. Ecol. 14, 859–867. (doi:10.1111/j.1365-294X.2005.
02452.x)
Juan, C., Emerson, B. C., Oromi, P. & Hewitt, G. M. 2000
Colonization and diversification: towards a phylogeo-
graphic synthesis for the Canary Islands. Trends Ecol. Evol.
15, 104–109. (doi:10.1016/S0169-5347(99)01776-0)
Kizirian, D., Trager, A., Donnelly, M. A. & Wright, J. W.
2004 Evolution of Gala
´
pagos Island lava lizards (Iguania:
Tropiduridae: Microlophus). Mol. Phylogenet. Evol. 32,
761–769. (doi:10.1016/j.ympev.2004.04.004)
Review. Diversification of Gala
´
pagos terrestrial fauna C. E. Parent et al. 3359
Phil. Trans. R. Soc. B (2008)

Kleindorfer, S., Chapman, T. W., Winkler, H. & Sulloway,
F. J. 2006 Adaptive divergence in contiguous populations
of Darwin’s small ground finch (Geospiza fuliginosa). Evol.
Ecol. Res. 8, 357–372.
Lack, D. 1947 Darwin’s finches. Cambridge, UK: Cambridge
University Press.
Lopez, T. J., Hauselman, E. D., Maxson, L. R. & Wright,
J. W. 1992 Preliminary analysis of phylogenetic relation-
ships among Gala
´
pagos Island lizards of the genus
Tropidurus. Amphibia-Reptilia 13, 327–339. (doi:10.1163/
156853892X00030)
Lundh, J. P. 2001 The Gala
´
pagos: a brief history. See http://
www.lundh.no/jacob/galapagos/pg05.htm.
McMullen, C. K. 1999 Flowering plants of the Gala
´
pagos.
Ithaca, NY: Cornell University Press.
Milinkovitch, M. C., Monteyne, D., Gibbs, J. P., Fritts, T. H.,
Tapia, W., Snell, H. L., Tiedemann, R., Caccone, A. &
Powell, J. R. 2004 Genetic analysis of a successful
repatriation programme: giant Gala
´
pagos tortoises. Proc.
R. Soc. B 271, 341–345. (doi:10.1098/rspb.2003.2607)
Milinkovitch, M. C., Monteyne, D., Russello, M. A., Gibbs,
J. P., Snell, H. L., Tapia, W., Marquez, C., Caccone, A. &
Powell, J. R. 2007 Giant Gala
´
pagos tortoises: molecular
genetic analysis reveals contamination in a repatriation
program of an endangered taxon. BMC Ecol. 7,2.(doi:10.
1186/1472-6785-7-2)
Naumann, T. & Geist, D. J. 2000 Physical volcanology and
structural development of Cerro Azul Volcano, Isabela
Island, Gala
´
pagos: implications for the development of
Gala
´
pagos-type shield volcanoes. Bull. Volcanol. 61,
497–514.
Nordlie, B. E. 1973 Morphology and structure of western
Gala
´
pagos volcanos and a model for their origin. Geol. Soc.
Am. Bull. 84, 2931–2955. (doi:10.1130/0016-7606(1973)
84!2931:MASOTWO2.0.CO;2)
Parent, C. E. & Crespi, B. J. 2006 Sequential colonization
and diversification of Gala
´
pagos endemic land snail
genus Bulimulus (Gastropoda, Stylommatophora). Evo-
lution 60, 2311–2328. (doi:10.1111/j.0014-3820.2006.
tb01867.x)
Peck, S. B. 1990 Eyeless arthropods of the Gala
´
pagos-
Islands, Ecuador—composition and origin of the crypto-
zoic fauna of a young, tropical, oceanic archipelago.
Biotropica 22, 366–381. (doi:10.2307/2388554)
Peck, S. B. 2001 Smaller orders of insects of the Gala
´
pagos
Islands, Ecuador: evolution, ecology, and diversity. Ottawa,
Canada: NRC Research Press.
Peck, S. B. 2006 The beetles of the Gala
´
pagos Islands, Ecuador:
evolution, ecology, and diversity (Insecta: Coleoptera).
Ottawa, Canada: NRC Research Press.
Peck, S. B. & Finston, T. L. 1993 Gala
´
pagos Islands
troglobites: the questions of tropical troglobites, parapatric
distributions with eyed-sister-species, and their origin by
parapatric speciation. Me
´
m. Biospe
´
ol. 20, 19–37.
Petren, K., Grant, B. R. & Grant, P. R. 1999 A phylogeny of
Darwin’s finches based on microsatellite DNA length
variation. Proc. R. Soc. B 266, 321–329. (doi:10.1098/
rspb.1999.0641)
Petren, K., Grant, P. R., Grant, B. R. & Keller, L. F. 2005
Comparative landscape genetics and the adaptive radi-
ation of Darwin’s finches: the role of peripheral isolation.
Mol. Ecol. 14, 2943–2957. (doi:10.1111/j.1365-294X.
2005.02632.x)
Powell, J. R. & Caccone, A. 2006 A quick guide to Gala
´
pagos
tortoises. Curr. Biol. 16, R144–R145. (doi:10.1016/j.cub.
2006.02.050)
Powell, J. R. & Caccone, A. 2008 CSI tortoise: unraveling the
mystery of mysteries. Gala
´
pagos News 26, 8–9.
Price, J. P. & Clague, D. A. 2002 How old is the Hawaiian
biota? Geology and phylogeny suggest recent divergence.
Proc. R. Soc. B 269, 2429–2435. (doi:10.1098/rspb.2002.
2175)
Pritchard, P. C. H. 1996 The Gala
´
pagos tortoises: nomenclatural
and survival status. Lunenburg, MA: Chelonian Research
Foundation.
Rassmann, K. 1997 Evolutionary age of the Gala
´
pagos
iguanas predates the age of the present Gala
´
pagos Islands.
Mol. Phylogenet. Evol. 7, 158–172. (doi:10.1006/mpev.
1996.0386)
Rassmann, K., Tautz, D., Trillmich, F. & Gliddon, C. 1997
The microevolution of the Gala
´
pagos marine iguana
Amblyrhynchus cristatus assessed by nuclear and mito-
chondrial genetic analyses. Mol. Ecol. 6, 437–452. (doi:10.
1046/j.1365-294X.1997.00209.x)
Russello, M. A., Glaberman, S., Gibbs, J. P., Marquez, C.,
Powell, J. R. & Caccone, A. 2005 A cryptic taxon of
Gala
´
pagos tortoise in conservation peril. Biol. Lett. 1,
287–290. (doi:10.1098/rsbl.2005.0317)
Russello, M. A., Beheregaray, L. B., Gibbs, J. P., Fritts, T. H.,
Havill, N., Powell, J. R. & Caccone, A. 2007a Lonesome
George is not alone among Gala
´
pagos tortoises. Curr. Biol.
17, R317–R318. (doi:10.1016/j.cub.2007.03.002)
Russello, M. A., Hyseni, C., Gibbs, J. P., Cruz, S., Marquez, C.,
Tapia, W., Velensky, P., Powell, J. R. & Caccone, A. 2007b
Lineage identification of Gala
´
pagos tortoises in captivity
worldwide. Anim. Conserv. 10, 304–311. (doi:10.1111/
j.1469-1795.2007.00113.x)
Sato, A., Tichy, H., O’Huigin, C., Grant, P. R., Grant, B. R.
& Klein, J. 2001 On the origin of Darwin’s finches. Mol.
Biol. Evol. 18, 299–311.
Schluter, D. 2000 The ecology of adaptive radiation. Oxford,
UK: Oxford University Press.
Schmitz, P., Cibois, A. & Landry, B. 2007 Molecular
phylogeny and dating of an insular endemic moth
radiation inferred from mitochondrial and nuclear genes:
the genus Galagete (Lepidoptera: Austostichidae) of the
Gala
´
pagos Islands. Mol. Phylogenet. Evol. 45, 180–192.
(doi:10.1016/j.ympev.2007.05.010)
Sequeira, A. S., Lanteri, A. A., Scataglini, M. A.,
Confalonieri, V. A. & Farrell, B. D. 2000 Are flightless
Galapaganus weevils older than the Gala
´
pagos Islands they
inhabit? Heredity 85, 20–29. (doi:10.1046/j.1365-2540.
2000.00690.x)
Sequeira, A. S., Lanteri, A. A., Roque-Albelo, L.,
Bhattacharya, S. & Sijapati, M. 2008 Colonization history,
ecological shifts and diversification in the evolution of
endemic Gala
´
pagos weevils. Mol. Ecol. 17, 1089–1107.
(doi:10.1111/j.1365-294X.2007.03642.x)
Sequeira, A. S., Sijapati, M., Lanteri, A. A. & Roque
Albelo, L. 2008 Nuclear and mitochondrial sequences
confirm complex colonization patterns and clear species
boundaries for flightless weevils in the Gala
´
pagos archi-
pelago. Phil Trans. R. Soc. B 363, 3439–3451. (doi:10.
1098/rstb.2008.0109)
Smith, A. G. 1966 Land snails of the Gala
´
pagos. In The
Gala
´
pagos. Proc. symposia of the Gala
´
pagos International
Scientific Project, 1964 (ed. R. I. Bowman), pp. 240–251.
Berkeley, CA: University of California Press.
Snell, H. L., Snell, H. M. & Tracy, C. R. 1984 Variation
among populations of Gala
´
pagos land iguanas (Conolo-
phus): contrasts of phylogeny and ecology. Biol. J. Linn.
Soc. 21, 185–207. (doi:10.1111/j.1095-8312.1984.tb0
2061.x)
Snell, H. M., Stone, P. A. & Snell, H. L. 1996 A summary of
geographical characteristics of the Gala
´
pagos Islands.
J. Biogeogr. 23, 619–624. (doi:10.1111/j.1365-2699.
1996.tb00022.x)
3360 C. E. Parent et al. Review. Diversification of Gala
´
pagos terrestrial fauna
Phil. Trans. R. Soc. B (2008)

Steinfartz, S., Glaberman, S., Lanterbecq, D., Marquez, C.,
Rassmann, K. & Caccone, A. 2007 Genetic impact of a
severe El Nin
˜
o event on Gala
´
pagos marine iguanas
(Amblyrhynchus cristatus). PLoS One 2, e1285. (doi:10.
1371/journal.pone.0001285)
Swanson, F. J., Baitis, H. W., Lexa, J. & Dymond, J. 1974
Geology of Santiago, Ra
´
bida, amd Pinzo
´
n Islands,
Gala
´
pagos. Geol. Soc. Am. Bull. 85, 1803–1810. (doi:10.
1130/0016-7606(1974)85!1803:GOSRAPO2.0.CO;2)
Thomas, R. A. 1997 Gala
´
pagos terrestrial snakes: biogeo-
graphy and systematics. Herpetol. Nat. Hist. 5, 19–40.
Thornton, I. 1971 Darwin’s islands: a natural history of the
Gala
´
pagos. New York, NY: Natural History Press.
Tonnis, B., Grant, P. R., Grant, B. R. & Petren, K. 2005
Habitat selection and ecological speciation in Gala
´
pagos
warbler finches (Certhidea olivacea and Certhidea fusca). Proc.
R. Soc. B 272,819–826.(doi:10.1098/rspb.2004.3030)
van der Werff, H. 1979 Conservation and vegetation of the
Gala
´
pagos Islands. In Plants and islands (ed. D. Bramwell),
pp. 391–404. London, UK: Academic Press.
Wagner, W. L. & Funk, V. A. 1995 Hawaiian biogeography.
Washington, DC: Smithsonian Institution.
Watkins, G. & Cruz, F. 2007 Gala
´
pagos at risk: a socioeconomic
analysis of the situation in the archipelago. Puerto Ayora,
Province of Gala
´
pagos, Ecuador: Charles Darwin
Foundation.
Werner, R., Hoernle, K., van den Bogaard, P., Ranero, C. & von
Huene, R. 1999 Drowned 14-m.y.-old Gala
´
pagos archipe-
lago off the coast of Costa Rica: implications for tectonic
and evolutionary models. Geology 27,499–502.(doi:10.
1130/0091-7613(1999)027!0499:DMYOGPO2.3.CO;2)
White, W. M., McBirney, A. R. & Duncan, R. A. 1993
Petrology and geochemistry of the Gala
´
pagos Islands:
portrait of a pathological mantle plume. J. Geophys. Res
Solid Earth 98,19533–19563. (doi:10.1029/93JB02018)
Whiteman, N. K., Kimball, R. T. & Parker, P. G. 2007
Co-phylogeography and comparative population genetics
of the threatened Gala
´
pagos hawk and three ectoparasite
species: ecology shapes population histories within parasite
communities. Mol. Ecol. 16, 4759–4773. (doi:10.1111/
j.1365-294X.2007.03512.x)
Whitlock, M. C. & Phillips, P. C. 2000 The exquisite corpse:
a shifting view of the shifting balance. Trends Ecol. Evol. 15,
347–348. (doi:10.1016/S0169-5347(00)01930-3)
Whittaker, R. J., Triantis, K. A. & Ladle, R. J. 2008 A general
dynamic theory of oceanic island biogeography. J. Biogeogr.
35,977–994.(doi:10.1111/j.1365-2699.2008.01892.x)
Wiggins, I. L. & Porter, D. M. 1971 Flora of the Gala
´
pagos
Islands. Stanford, CA: Stanford University Press.
Wikelski, M., Foufopoulos, J., Vargas, H., Snell, H. 2004
Gala
´
pagos birds and diseases: invasive pathogens as
threats for island species. Ecol. Soc. 9, article no. 5. See
http://www.ecologyandsociety.org/vol9/iss1/art5.
Wright, J. W. 1983 The evolution and biogeography of the
lizards of the Gala
´
pagos archipelago: evolutionary genetics
of Phyllodactylus and Tropidurus populations. In Patterns of
evolution in Gala
´
pagos organisms (eds R. I. Bowman, M.
Berson & A. E. Leviton), pp. 123–155. San Francisco,
CA: American Association for the Advancement of
Science, Pacific Division.
Wyles, J. S. & Sarich, V. M. 1983 Are the Gala
´
pagos iguanas
older than the Gala
´
pagos? Molecular evolution and
colonization models for the archipelago. In Patterns of
Evolution in Gala
´
pagos Organisms (eds R. I. Bowman,
M. Berson & A. E. Levinton), pp. 177–185. San Francisco:
American Association for the Advancement of Science,
Pacific Division.
Review. Diversification of Gala
´
pagos terrestrial fauna C. E. Parent et al. 3361
Phil. Trans. R. Soc. B (2008)
